Les virus, parasites du systèmes d’expression des cellules
Historique : Pasteur et Chamberland montrent que des maladies sont transmises par un filtrat dont on avait éliminé toute bactérie.
Virus trop petit pour être arrêtés par les filtre en porcelaine très fins («bougie» de Chamberland : 0,2 micrometre)
1892: Iwanowski apporte la preuve que la mosaïque du tabac est due à un virus.
1929 : 1ere photo MET du VMT
Lwoff : un virus est un virus !
3 exemples sont étudiés : VMT (cristallisé en 1935), phage lambda (infectant la souche K12 d’E.Coli. Les bactéries sont lysogènes, les phages sont tempérés.) et le HIV ( HIV1 et HIV2, isolé par Montagnier et Barré-Sinoussi en 1982 (nobel 2008)
-
I)Les virus : une organisation simplifiée ne permettant pas l’expression de l’information génétique virale
-
-
1)Une ou plusieurs enveloppes protégeant un acide nucléique: une taille réduite
-
-
-
-
A)Virus nus
-
-
-
•Virus possèdent toujours au moins une enveloppe qui protège l’acide nucléique.
-
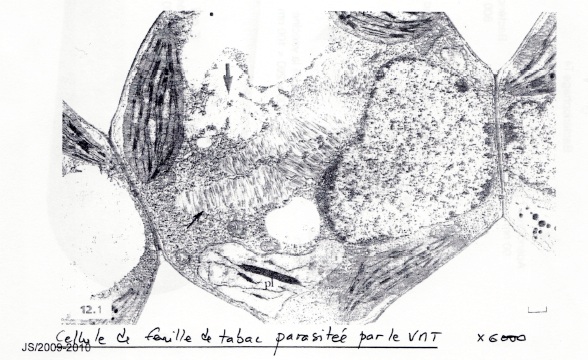
•Virus à nucléocapside à symétrie hélicoïdale : VMT
-
•Virus présent au centre sous forme d’amat. Le chloroplaste ne possède pas de granum, ce qui explique les tâches jaunes sur les feuilles.
Le VMT est formé par une seule enveloppe formé de 2130 proteines identique ➞ économie de matériel génétique pour fabriquer l’enveloppe. Le VMT mesure 17*300 nm. C’est un Virus ARN +.
-
-
•Virus à nucléocapside hélicoïdale et icosaédrique : phage λ
-
•

-
•Capside uniquement protéique formé de 2 types de glycoproteines ( gpE et gpD). Les glycoproteines s’assemblent en capsomères qui eux-même s’assemblent par 20 en triangle ➞ 20 capsomère ➞ capside.
-
-
-
B)Virus enveloppés
-
-
-
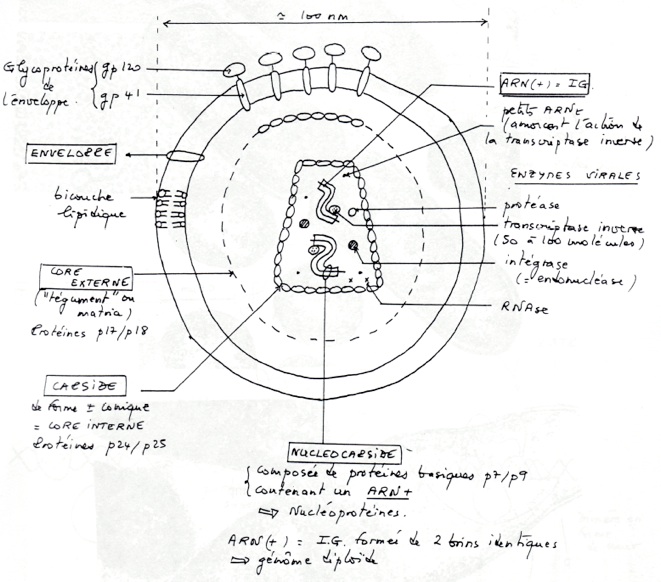
•Virus de l’Immunodéficience Humaine ( HIV )
-
-
2)Le nombre limité d’enzymes et de protéines régulatrices, une absence de hyaloplasme, d’organites et de métabolisme
-
-
-
•VMT : présence d’une enzyme : lysozyme.
-
•Phage lambda : possède aussi un lyzosyme
-
•HIV : enzyme : transcriptase reverse ( TR = 50 à 100 molécules), une protéase et une intégrase. Il contient 4 types de protéines régulatrices. Il contient aussi de l’ARNtLYS qui est une amorce de la TR.
-
•Les virus sont dépourvus de tout hyaloplasme et de tout organite. S’il peut y avoir des enzymes, aucune n’a de fonction métabolique.
-
-
3)Un génome réduit sous forme d’ADN ou d’ARN
-
-
-
•La taille moyenne d’un génome virale : 5 à 10 kb.
-
•VMT : génome formé d’un ARN monocaténaire de 7kb, pourvue de 4 gènes
-
•Phage lambda : ADN double brin de 48,5 kb, pourvue de 50 gènes codant des polypeptides organises en froupes fonctionnels (gènes précoces . Gènes tardifs).
-
•HIV ARN(+) de 9,2kb. Le génome code pour 15 protéines principales. Les séquences codantes organisées en 3 régons principales indispensables au cycles du Virus :
-
•GAG : proteines de la capside
-
•POL : enzymes
-
•ENV : protéines d’enveloppe
-

-
•La transcription va être contrôler par des protéines virale ET par des protéines cellulaire ! Il y a détournement des protéines cellulaires.
-
•La transcription va faire intervenir l’ARN polymérase II de la cellule. Les ARN formés vont être traduits et vont formé des poly-proteines qui sont des précurseurs. Ces poly-protéines vont subir une maturation post-traductionnelle visant à l’élaboration de protéine de l’envelloppe nucléocapside, des protéase, des TI, des intégrase et des protéines d’enveloppe.
-
•Chez les virus l’information est maximal dans une espace minimal : Réduction des génomes compensés par :
-
•Utilisation de plusieurs cadres de lecture ( gènes chevauchants)
-
•Slipage : glissement des ribosomes qui permet de sauter d’un cadre de lecture à l’autre
-
•Epissage différentiel ( HIV : 20 transcrits différents à partir du même messager)
-
•Maturation protéolytique : poly-protéine qui est clivée après traduction. Parfois clivage différentiel en fonction du moment du cycle du virus.
-
•Séquences intergéniques rares, cours ou inexistantes
-
•Séquences régulatrices communes à plusieurs gènes : expression des gènes en batteries (gènes précoces, gènes tardifs)
-
•Quand le virus a de l’ADN à double brin : utilisation des 2 brins qui sont informatifs.
-
-
II)Les virus pénètrent dans la cellule hôte et s’y multiplient en utilisant le système d’expression de celle-ci
-
-
1)Adsorption et pénétration dans l'hôte grâce au système de signalisation cellulaire
-
-
-
•L’adsorption des virions résulte d’un détournement de l’activité de reconnaissance cellulaire
-
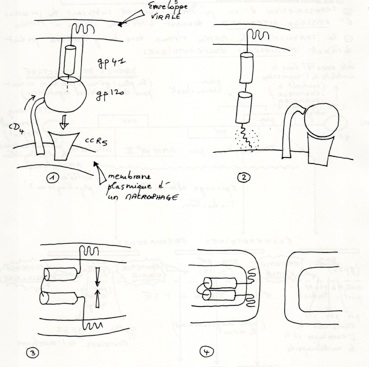
•Cas du HIV :
-
•Récepteurs CD4 ( CD = classe de différenciation) (sur macrophage, monocyte, LT4)
-
•Co-récepteurs CCR5 (=récépteur de chimiokine (ex interleukine)) (sur macrophages) et CXCR4 (sur lymphocytes T4)
-
-
•GP120 reconnaît le récepteur CD4, la molécule GP41 par GP120, et lors du contact GP120 et CD4, la conformation change ce qui provoque une interaction avec le récepteur CCR5. Une fois ceci fait, la molécule GP41 est libéré, et sa séquence hydrophobe terminale s’introduit dans l'enveloppe de la cellule. Une fois fait il y a rapprochement des 2 membranes et leur fusion.
-
•Cas du phage lambda : Se fixe sur le transporteur de maltose = protéine de la familles des porines codée par le gène lam B inductible par le maltose.
-
•Cas du VMT : Le virus pénètre par une blessure ou par des insectes vecteurs (pucerons). Il y a une absence de reconnaissance.
-
-
2)Des cycles ou alterne des phases de multiplication et des phase d’arret
-
-
-
-
A)Diversité des cycles viraux
-
-
-
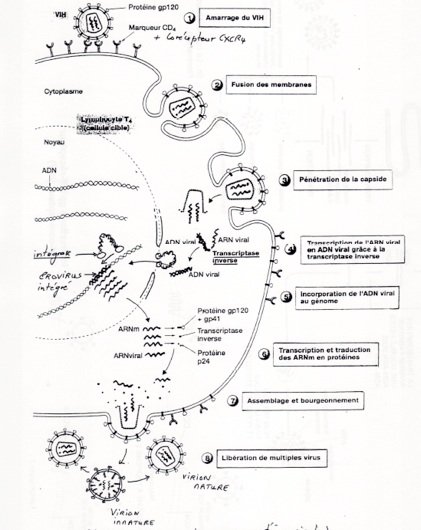
•Cycle du VIH :
-
•Information génétique est réduite : utilisation de l’information génétique de l'hôte pour produire ses protéines et ses ARN viraux.
-
•Multiplication par utilisation de la machinerie réplicative de l’hôte
-
•Pas de métabolisme : utilisation du métabolisme de l’hôte
-
•Utilisation des organites de l’hôte.
-
•Phase d’arret : virions, forme quiescente du HIV.
-

-
•Etape conduisant à l’assemblage et au bourgeonnement des virions :

-
•Cycle du phage lambda :
-
•
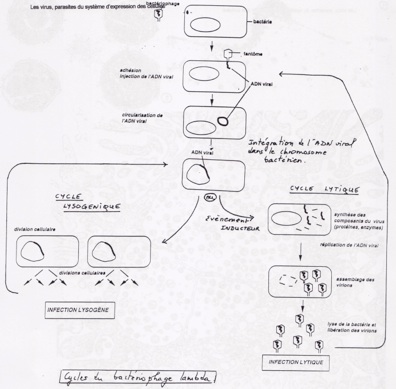
-
•Lors d’une culture normale d’E.Coli, on voie quelque plage de lyse, par contre, en présence d’UV, la lyse se généralise. L’intervention de facteurs pouvant conduire la mort cellule conduise a à un cycle lytique.
-
•Cycle lytique (45min à 37°c) . Epression séquentielle des gènes par blocs fonctionnels
-
•Phase précoce production des enzymes nécessaires à la réplication et a blocage de la traduction des ARNm cellulaire.
-
•Phase tardive début de réplication et synthèse des protéines de structure
-
•Cycle lysogénique : intégration site spécifique de l’ADN phagique dans le génome bactérien : prophage.
-
•Cycle du VMT :


-
•Le VMT se conserve dans les semence, débris végétaux, terreaux... Il peut se transmettre par les pratiques culturales (repiquage, tailles des têtes...)
-
•Particularité des cycles viraux :
-
•Phases d’arret :
-
•Sous formes de virions hors des cellules
-
•Sous formes quiescente (HIV)
-
•Sous formes lysogènes (phage lambda)
-
•Phase de multiplication toujours à l’intérieur de la cellule hôte.
-
-
-
B)Le passage du cyle productif aux phases d’arrêt
-
-
-
•Sortie des particules virales
-
•Lsozyme (=endolysine) synthétisé en fin de phase tardive chez les phages
-
•HIV par bourgeonnement
-
•Passage au cycle lytique : cas du phage lambda.
-
III)Les virus parasitent l’information génétique de l’hôte
-
-
1)Les virus se servent de l’information génétique pour multiplier leur ADN (intégration)
-
-
-
•Cf précédemment.
-
-
2)Les virus mettent le génome de l’hôte hors circuit ou modifient son expression et expriment leurs gènes
-
-
-
•Dans le cas du phage : Dans le cas du HIV :
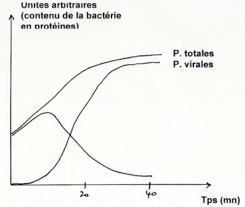
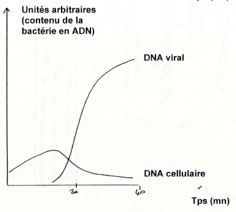
-
•L’ADN des bactérie est détruit par le phage. Il inhibe la synthèse des ARN cellulaire, bloque la traduction des ARNm bactériens par des protéines virales, inhibe la réplication de l’ADN cellulaire et la division cellulaire et enfin fragmente l’ADN de l'hôte par endonucléase virales (cas du cycle lytique).
-
-
3)Les virus peuvent influencer l’état immunitaire de la cellule hôte
-
-

IV) Les virus utilisent les ressources et la machinerie métabolique de l’hôte (pour exprimer leur information génétique)
-
•Utilisation d’organites cellulaires (pour le HIV, on a pus montrer que les microtubules cellulaire servait à acheminer les vésicules, les ribosomes pour la traduction)
-
•Utilisation de métabolites (utilisation des nucléotides, utilisation des AA, utilisation d’oses et d’ATP...)
-
•Utilisation de protéines cellulaire, principalement d’enzymes
-
•Kinases : décapsidation
-
•ARN polymérase qui sont dépendant d’ARN cellulaire : réplication de l’ARN viral
-
•ARN polymérase II cellulaire assurant la transcription des gènes précoces (phage lambda)
-
•Protéases cellulaire clivant ( en GP120 et GP41) le précurseur d’enveloppe du HIV
-
•L’amorce de la TR (HIV) est un ARNtLYS provenant de l’hôte
-
•Le VIH dépend de plus de 200 facteurs cellulaires
-
-
V)Effet des virus ur les cellules parasites
-
-
1)Les virus peuvent procoquer la mort des cellules parasités
-
-
-
•Mort par nécrose (différent de l’apoptose). La cellule infecté est dégénéré et amène a relacher les virus dans le milieu.
-
•Mort par lyse cellulaire.
-
•Destruction de la cellule infecté par le système immunitaire de l’organisme (HIV)
-
•Déclenchement de l’apoptose (HIV)
-
•Modification lié à l’apoptose :
-
•Boursouflement de a membrane plasmique
-
•Fragmentation de la cellule en vésicule = corps apoptotique entouré d’une membrane qui vont ensuite etre détruit par phagocytose.
-
•Condensation de la chromatine et diminution du volume cellulaire ➞ ADN fragmenté.
-
-
-
2)Evolution tumorale des cellules parasitées
-
-
-
•Le HIV peut être oncogène de façon indirecte en diminuant les défenses immunitaires antitumorales de l’organisme :
-
•Sarcome de Kaposi est fréquent chez les sidéens : fait d’une co-infection HIV-virus de type herpès.
-
•Un certains nombre de virus peut rester latent pendant plusieurs années et sans provoquer de symptômes. Certains facteurs externe peuvent entraîner la reprise du cycle virale.
CONCLUSION :
Virus = simplicité d’organisation qui survivent par expression de l’hôte. Ce ne sont pas des cellules par 3 caractères fondamentaux (absences de métabolisme, absence d’organites, incapacité de se multiplier hors des cellules). Dans un virus il y a toujours qu’un seul type d’information génétique (soit ADN, soit ARN).
Virus = être vivant ? N’a pas de sens en tant que tel, car hors des cellules il est inertes, mais semble vivant lorsqu’il est dans une cellule.
Origine des virus ? 2 théories se valent :
ils proviendraient d’une simplification de micro-organisme plus complexe. De petites bactéries qui vivaient en symbiose avec des cellules (probablement valide pour les gros virus : variole, vaccine)
Autonomisation progressive de certaines gènes cellulaires, en particuliers ceux codant pour les enzyme de la réplication ( les rétrovirus ont probablement cette origine)
