Le mécanisme de la conversion de l’énergie lumineuse en énergie chimique
INTRODUCTION: La photosynthèse permet de fabriquer les briques élémentaires des éléments indispensables à la vie. La photosynthèse est à l’origine de l’essentielle de l’oxygène sur terre. Elle est à l’origine des combustibles fossiles que nous utilisons encore aujourd’hui. Les chloroplastes sont des micro-centrales solaire convertissant l’énergie lumineuse en énergie chimique, ce qui pourrait être important pour l’avenir s’ils étaient entièrement compris.
-
I)Quelques jalons historiques dans la découvertes du mécanisme de la photosynthèse
-
•Priestley en 1771-1777 découvre qu’il y a rejet d’un gaz par une plante verte qui entretient la vie.
-
•Ingen-houz (1779) découvre que lumière et feuilles vertes sont indispensables. Il y a rejet d’oxygène seulement à la lumière.
-
•Senebier 1782 : les plantes absorbent du CO₂ en même temps qu’elles rejettent du dioxygène.
-
•De Saussure 1804 : les plantes absorbent de l’eau nécessaire à la photosynthèse.
-
•Equation de la photosynthèse :
6 CO₂ + 6 H₂O ➞ C6H12O6 + 6 O₂ -
•Mayer en 1842 suggère l'existence d’une conversion d’énergie lumineuse en énergie chimique.
-
•Sachs en 1864 montre que les plantes synthétisent de l’amidon dans leurs feuilles verts.
-
•Van Niel en 1931 montre que les réactions photosynthétiques sont des réactions d’oxydo-réduction.
-
•Démonstration du rôle de l’eau dans la production du dioxygène par Rubben et Kamen (1941), ils utilisent de l’eau marqué avec de l’oxygène lourd : 18O
Résultats : la plante ayant reçus l’eau marquée avec 18O₂ ont produit de l’O₂ marqué. L’O₂ n’a donc pas pour origine le CO₂ mais l’eau qu’absorbe la plante par ses racines. -
•On a montré ensuite qu’il existait une phase photochimique et une phase chimique. Les produits formés (glucides) peuvent être révélés par radiochromatographie.
-
II)Mise en évidence de l'existence de 2 phases distinctes dans la photosynthèse
-
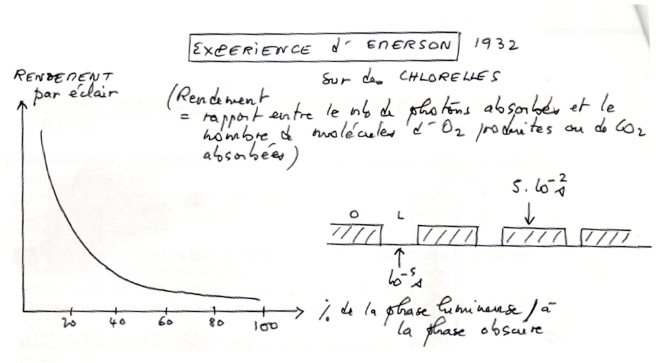
•Expérience d’Emerson en 1932:
-
•Résultat :
Le rendement par éclair est une fonction décroissante de l’importance de la phase lumineuse.
Rendement est maximum pour des éclairs de 10-5 seconde séparés par des intervalles d’obscurité de 5.10-2 seconde
-
•Interprétation :
La lumière est d’autant mieux utilisées que les produits formés très rapidement à la lumière sont utilisés au cours de la phase chimique.
Autrement dit, les réactions photochimiques nécessitent des substrats en quantité limite (ADP, NADP+) qui doivent être régénérés au cours de réactions non photochimiques :
une excitation lumineuse avant cette régénération est inefficace.

-
•La production d’O₂ peut se produire sans réduction de CO₂ , cela confirme que l’O₂ vient de l’H₂O et non du CO₂
-
•Des chloroplastes isolés peuvent réaliser une partie des réactions de la photosynthèse.
-
•Cela révèle que la première phase de la photosynthèse est le transfert d’électrons de l‘eau à une autre substance contre un potentiel chimique
-
•Photo-oxydation de l’eau ➞ conversion d’énergie lumineuse en énergie chimique.
On voit que l’incorporation de CO₂ se prolonge à l’obscurité.
-
•Il est possible de montrer ainsi qu’il existe 2 phases distinctes de la photosynthèse:
-
-
• Phase photochimique nécessitant la lumière.
-
• Phase chimique ne nécessitant pas la lumière (mais se déroulant à la lumière et pouvant se prolonger quelque instant à l’obscurité.
-
-
-
III)La collecte de l’énergie lumineuse et son transfert au centre réactionnel
-
-
1)Les pigments chlorophylien dans a photosynthèse
-
-
A/ Mise en évidence du rôle des pigments chlorophylliens dans la photosynthèse
-
•Expérience triviale avec des caches et des feuilles.
B/ Extraction et nature des pigments chlorophylliens
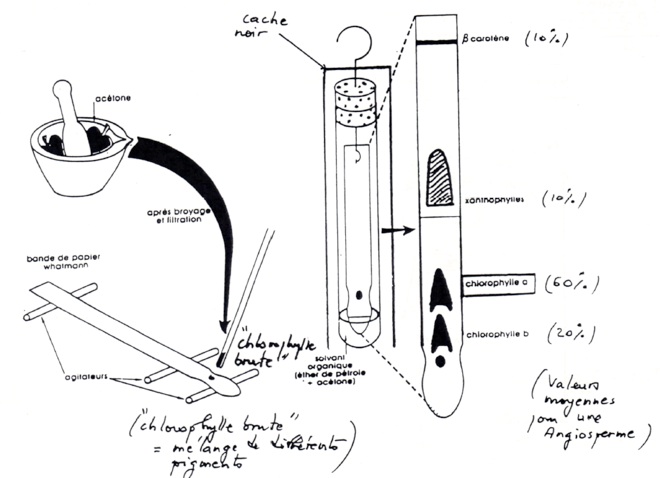
-
•Chlorophylles et pigments accessoires sont des molécules excitables par des photons de lumière visible = vecteurs d’une énergie qui permet la transition d’un électron π vers une orbitale plus externe.
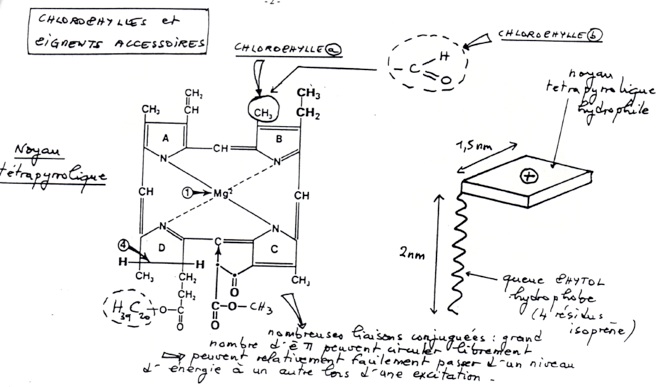
-
•Il existe d’autres pigments, comme la carotènes et la xanthophylles. Ils permettent d’augmenter le nombre des longueurs d’ondes qui sont absorbées. Ces pigments sont associés par des liaisons covalentes à des polypeptides (chlorophylles comprise)
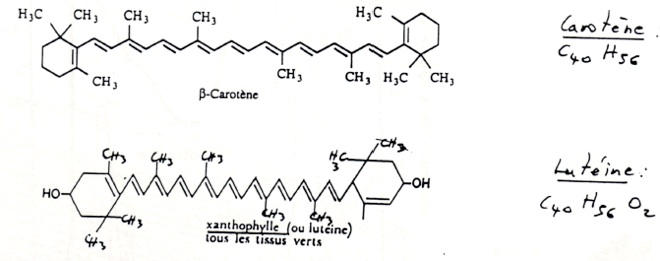
C/ Mise en évidence des fonctions des pigments chlorophylliens
-
•Mesure des spectres d’absorption et mesure du spectre d’action :
-
•
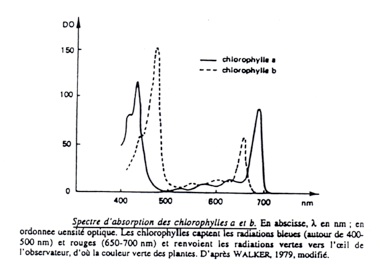
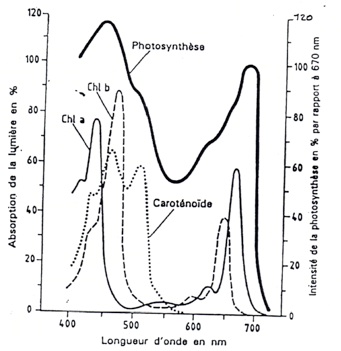
-
-
2)Les chloroplastes et le thylakoïde : des structures adaptées à la collecte de l’énergie lumineuse
-
-
A/ Les chloroplastes
-
•La forme, la dimension et le nombre des chloroplastes est variables selon les espèces. (forme lenticulaire chez les végétaux supérieur, mais hélicoïdale chez la Spirogyre, en coupe chez les Chlamydomonas, étoilé chez les Zygnema...).
De 20 à 60 dans les cellules d’épinard (en général 20 à 40 par cellule). Ils sont mobiles dans le cytoplasme, entraînés pas des mouvements de cyclose. -
•Organites compartimentés composés de 3 compartiments distincts.
-
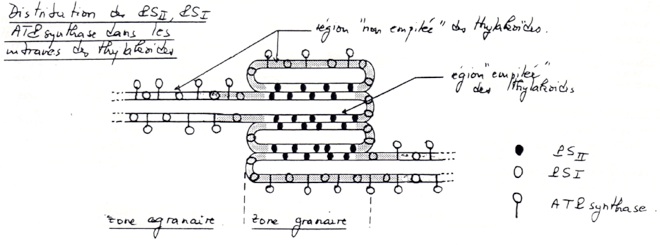
•Grande surface, abondance de membrane sous forme de thylakoïdes contenant les photosystèmes et les chaînes de transporteurs d’électrons.
-
•Possibilité d’établissement d’un gradient de protons
-
•Ségrégation possible de réaction. Petit compartiment confinés favorisant les rencontre ES.
-
•Echanges sélectifs possibles avec le hyaloplasme.
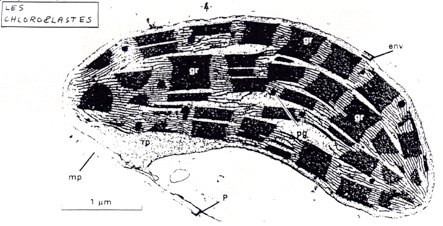
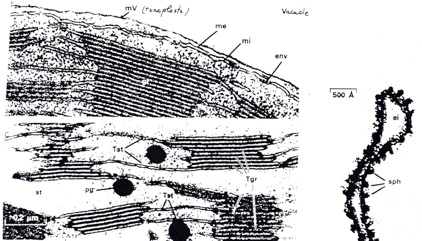
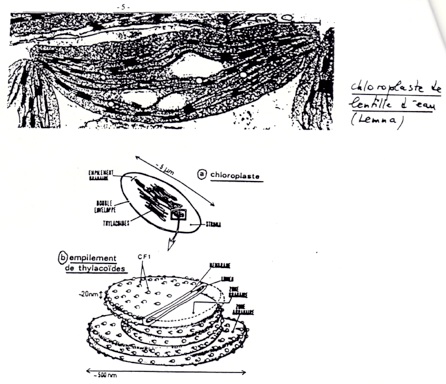
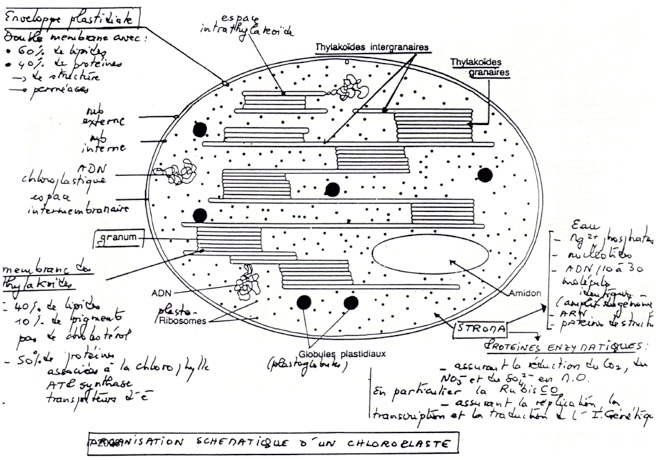
B/ Organisation et composition des thylakoides.
-
•Obtention et analyse de sous fractions chloroplastiques:
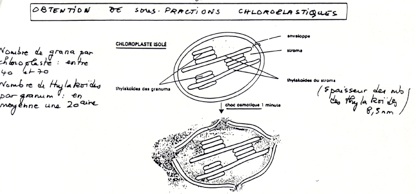

-
•Il y a une opposition entre les 2 membranes :
-
•Membrane externe : contient des porines (perméabilité à des solutés jusqu’à 10 000 Da)
-
•Membrane interne : relativement perméable (présence de transporteurs) : perméabilité sélective.
-
•Membrane des thylakoides
-
•50% de protéines
-
•40% de lipides : richesse en galactolipides (40% des lipides totaux. Contiennent des acides gras avec plusieurs doubles liaisons). Membrane fluide donc diffusion facilités des complexes protéiques intervenant dans la photosynthèse.
-
•10% de pigments.
-
-
-
3)les effets des photons sur les pigments chlorophylliens, conséquences possibles
-
-
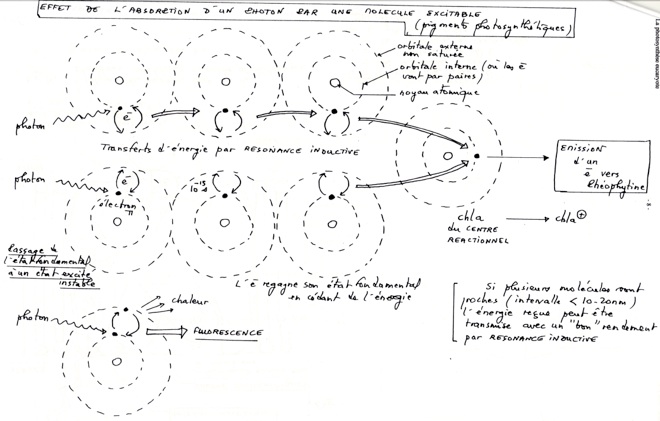
A/ Passage de l’état fondamental à l’état excité (exciton)
-
•Les pigmentes absorbent des photons et passent à l’état excités (réactif) instable. L’état excité ne dure qu’environ 10-9 seconde (Des atomes donnés ne peuvent être portés à l’état excité que par certaines longueurs d’ondes.)
B/ Conversion de l’état excité à l’état fondamental
-
•L’électron sur l’orbitale externe peut revenir a son orbitale d’origine selon plusieurs modalités :
-
•Emission de chaleur.
-
•Libération d’un quantum d’énergie plus faible. Comme il y a perte d’énergie, les photons émis ont une énergie moindre c’est à dire une longueur d’onde plus grande que ceux absorbés. Fluorescence infra rouge et rouge (in vivo = 3% de la lumière absorbé)
-
•Transfert d’excitation par résonance inductive dans le sens des longueurs d’ondes croissantes. Conduit à un maximum de radiations absorbées pas la chlorophylle A. Ce transfert se fait jusqu’au dimère de chlorophylle A, la convergence des excitation par résonance amène celle-ci à l’expulsion d’un électron, et à sa photo-oxydation.
-
C/ Modification chimique du pigment = photo-oxydation (=conversion d’énergie photonique/ énergie chimique)
-
•L’électron sur l’orbitale externe est transféré à une autre molécule = conversion externe.
-
-
4)Collecte des photons et transfert de l’énergie au sein des photosystèmes
-
-
A/ Mise en évidence de l’existence de 2 photosystèmes : expérience d’EMERSON
-
•Globalement, les faibles longueurs d’ondes «aident» les longueurs d’ondes plus élevées. De ça, Emerson émet l’idée qu’il existe 2 systèmes d’énergie lumineuse qu’il appellera photosystème, l’un actif jusqu'à 680nm, et un autre actif jusqu'à 700nm , et ces 2 photosystème ne sont pas indépendant.
B/ Organisation et fonctionnement des photosystèmes
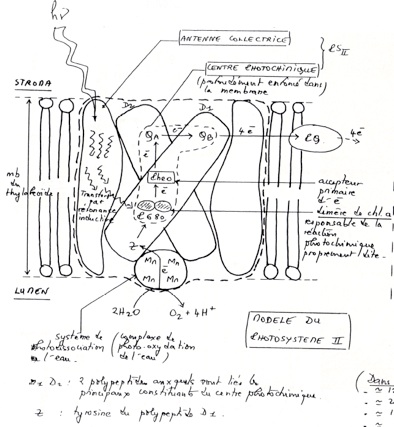
Les électrons circulent dans une chaînes de transporteur présentes dans la membrane des thylakoïdes.
-
III)Les transferts d’électron d’un réducteur faible (l’eau) à un réducteur puissant le NADPH,H+
-
-
1)Etape du transfert et couplage mis en jeu
-
-
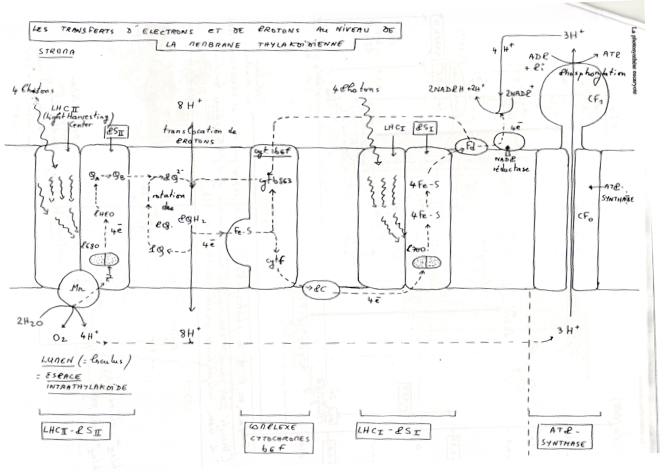
-
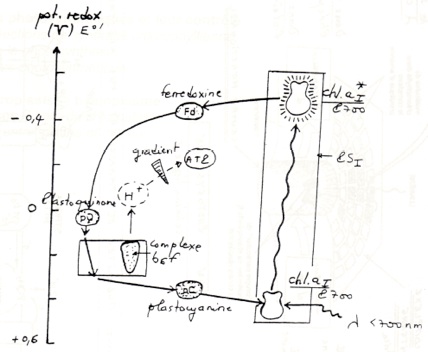
•Dans le photosystème II, le trou électronique créé par le départ d’électron vers la phéophytine est comblé par des électrons provenant de la photo-oxydation de H₂O . Ces 4 électrons sont transmis par un système de décomposition de l’eau lié à P680 (= un système contenant 4 atomes de Mn et une TYR du polypeptide D1).
-
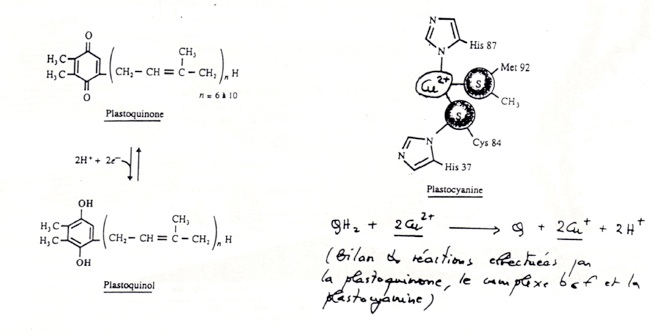
•Dans la phéophytine, les électrons passent à des quinones A et B du photosystème II puis à une petite molécule mobile, la plastoquinone.
La plastoquinone est un transporteur d’électrons et de H+. Elle transfère ses électrons à un complexe B6F. Le complexe B6F accepte les électrons mais pas les protons : ceux-ci sont transloqués dans le lumen. -
•Complexe B6F : (équivalent du complexe BC1 de la mitochondrie).
Plusieurs sous unités : cytochrome B6, f, protéine fer-souffre.
C’est une site de translocation de protons.
Les électrons sont ensuite transférés à la plastocyanine. -
•La plastocyanine : petite molécule mobile ( contenant du Cu2+ comme cofacteur) située sur la membrane coté lumen.
Transfert liés à la chlorophylle A, du photosystème I dans le trou électronique laissé après sont oxydation. -
•De la chlorophylle A, les électrons passent de A0 (une chlorophylle A) puis à A1 (une phyloquinone), puis à 3 centre Fe-S faisant partie du centre réactionnel du photosystème I.
Les électrons sont ensuite transférés à une petite protéine Fe-S : la ferredoxyne. -
•La ferredoxyne transmet les électrons à l’accepteur finale, le NADP+.
La réduction du NADP+ est catalysée par le complexe ferredoxine-NADP réductase, une volumineuse enzyme.
-
-
2)Bilan énergétique
-
-
-
•Depuis l’eau : potentiel redox O2/H2O = +0,82V, jusqu’au NADP: potentiel redox NADP+/NADPH2 = -0,32V, 4 électron ont été transférés à partir de l’eau grâce à l’énergie lumineuse : ils ont subi une E°’ de 1,13V d’où : G°’ = nF . E°’ = 4,9.96699.1,13.10-3

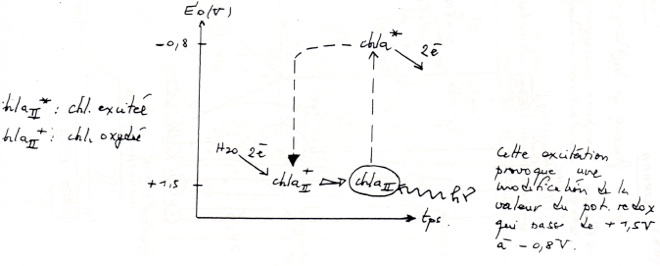
-
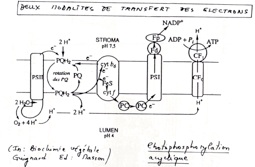
IV)La photophosphorylation acyclique
-
-
1)Mise en évidence expérimentale du rôle d’un gradient de protons dans les phosphorylations
-
-
-
•Expérience de Jagendorf en 1966 qui met en évidence le gradient de proton dans le processus de phosphorylation. Il a utilisé des thylakoïdes isolés et à créer artificiellement un gradient de protons à partir duquel il y observe les différents effets.
-
•On tire de cette expérience que le gradient de protons est indispensable à la synthèse d’ATP. De plus, on a montré que les têtes des ATPsynthases sont indispensables. Un protonophore, les DNP (Dinitrophénol) un composé qui rend perméable la membrane au proton empêche la formation d’un gradient de protons, et on n’observe pas de formation d’ATP.
-
-
2)Les ATPsynthases, agents du couplage osmo-chimique
-
-
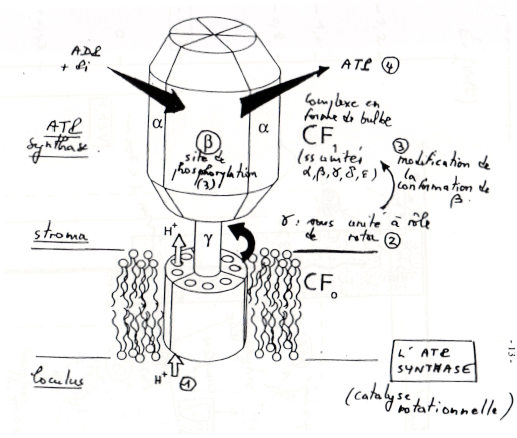
-
•Le complexe moléculaire comprend une partie enchâssée dans la membrane avec de nombreux polypeptides. Elle contient les canaux à protons. Une seconde partie fixe et ne pouvant tourner est reliée par une troisième qui la relie à la partie membranaire, rotatoire.
Fonctionnement : Protonation d’un aspartate et déprotonation lors de la traversé des canaux. Cette traversée fait tourner la partie γ centrale, et cette rotation change la conformation des sous unités β qui lancent la phosphorylation de l’ADP en ATP.
➞ On a une catalyse rotatoire.

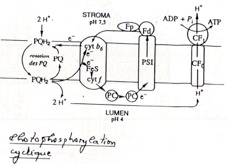
V) Une photophosphorylation qui peut être cyclique
-
•Il y a production d’ATP sans production de NADPH,H+.
-
•Mise en évidence grâce au DCMU (= dichlorophénylméthyl urée), qui est un poison. On observe avec ce poison qu’il y a toujours synthèse d’ATP mais plus de synthèse de NADPH,H+. On simule donc un mécanisme qui est possible dans les cellules végétales.
-
•Rôle du DCMU ? Il bloque le passage des électrons entre la Chlorophylle A et la plastoquinone. Il met donc hors circuit le photosystème II : Il découple donc les 2 photosystèmes. En conséquence, les électrons sortant de la Chlorophylle A ne vont pas à la ferredoxyne mais ils retournent au niveau de la plastoquinone, ils forment ainsi un circuit fermé.
-
•Ce système est mis en oeuvre lorsque la teneur en NADPH,H+ est suffisante et toute l’énergie solaire récoltée est utilisée pour la fabrication d’ATP. Cette voie de photophosphorylation cyclique permet d’adapter le rapport ATP/NADPH,H+ au besoin de la cellule. Il s’agit là d’une forme de régulation.